Исходя из физиологических представлений, нервный «центр» может располагаться на разных уровнях ЦНС и участвовать в регуляции какой-либо фиpиологической функции (дыхание, пищеварение и т.д.) или в совершении какого-либо рефлекса.
К функциональным свойствам рефлекторных центров относятся: иррадиация
возбуждения; конвергенция
и дивергенция
; суммирование; синаптическое облегчение и окклюзия; трансформация ритма, реверберация возбуждения; тоническое состояние центров, их быстрая утомляемость, большая чувствительность к недостатку кислорода и к действию некоторых ядов.
Иррадиация возбуждения
Активное распространение возбуждения в ЦНС, особенно при сильном и длительном раздражении, получило название иррадиации. Возможность иррадиации в ЦНС обусловлена наличием в ней многочисленных ответвлений отростков (аксонов, дендритов) нервных клеток и цепей интернейронов, которые соединяют между собой различные нервные центры (благодаря этому возбуждение распространяется определенными путями и с определенной последовательностью). Важную роль в иррадиации возбуждения в структурах мозга играет .Усиление раздражения или повышение возбудимости ЦНС сопровождается усилением иррадиации возбуждения в ней. Тормозные нейроны и синапсы препятствуют иррадиации возбуждения или ограничивают ее. При введении стрихнина, блокирующего постсинаптическое торможение, возникает сильное возбуждение ЦНС, которое сопровождается судорогами всех скелетных мышц. Иррадиация может стать патологической в связи с возникновением сильного очага возбуждения и с изменением свойств нервной ткани, усиливает распространение возбуждения. Это бывает при эпилепсии .
Конвергенция возбуждения
На каждом из нейронов ЦНС конвергирует (сходятся) различные афферентные волокна. Таких афферентных входов для большинства нейронов много десятков и даже тысяч. Так, на мотонейронах заканчиваются в среднем 6000 коллатералей аксонов, которые поступают от периферических рецепторов и различных структур мозга, образуя возбуждающие и тормозные синапсы. Это такое универсальное явление, можно говорить о принципе конвергенции в нейронах и их связях. Благодаря этому явлению в один и тот же нейрон одновременно поступают многочисленные и разнообразные потоки возбуждений, которые затем подлежат сложной обработке и перекодируются и формируются в единое возбуждение - аксонноу, что идет к следующему звену нервной сетки. Конвергенция возбуждения на нейроне является универсальным фактором его интегративной деятельности.Различают мультисенсорную, мультибиологическую и сенсорно-биологическую формы конвергенции. В первом случае на нейрон поступают сигналы различной сенсорной модальности (зрительные, слуховые, болевые и др.), во втором - потоки возбуждений различной биологической модальности (пищевые, половые и др.), в третьем - сигнализация (зрительная, пищевая) и другие.
Дивергенция возбуждения
Дивергенция (расхождение) возбуждения - способность одиночного нейрона устанавливать в многочисленных синаптических связях с различными нервными клетками. Например, афферентные волокна периферических рецепторов, входя в спинной мозг в составе задних корешков, дальше разветвляются на многочисленные коллатерали, которые идут к спинальным нейронам. Благодаря дивергенции одна и та же нервная клетка может принимать участие в организации различных реакций и контролировать большое количество нейронов. Одновременно каждый нейрон может обеспечивать широкое перераспределение импульсов, что ведет к иррадиации возбуждения. Конвергенция и дивергенция взаимно связаны.Реверберация возбуждения
Циркуляция возбуждения замкнутыми нейронами и их цепями в ЦНС называется реверберацией. Возбуждение одного из нейронов, входящих в эту цепь, передается на другой (или другие), а коллатералям аксонов снова возвращается к нервной клетки и т.д.Реверберация возбуждения наблюдается в так называемом рефлекторном последействии, когда рефлекторный акт заканчивается не сразу после прекращения, а через некоторый (иногда длительный) период, а также играет определенную роль в механизмах кратковременной (оперативной) памяти. Сюда же относится корково-подкорковая реверберация, которая играет важную роль в высшей нервной деятельности (поведении) человека и животных.
Тонус нервных центров
Многие центры, т.е. нейронов, которые их составляют, постоянно генерируют нервные импульсы. Они поступают от эффекторов, что свидетельствует о существовании некоторого постоянного тонического возбуждения, т.е. тонуса нервных центров.Указанное свойство нервного центра проще рассмотреть на примере объединения мотонейронов (мотонейронного пула).
При раздражении афферентного мышечного нерва надпороговым одиночным стимулом мотонейрона, иннервирующего соответствующие мышцы, возникает моносинаптический ВПСП. В зависимости от числа синаптических контактов и уровня поляризации часть мотонейронов деполяризуется до порогового уровня, и в них происходит импульсивный разряд. Эти мотонейроны составляют так называемую зону разряда. Вторая (обычно значительно большая) часть мотонейронов этого пула не достигает критического уровня деполяризации и не разряжается, но на время развития ВПСП, как правило, увеличивается возбудимость этих «молчаливых» нейронов. Эти нейроны составляют так называемую подпороговую зону нервного центра.
Подпороговая зона увеличивается при усилении афферентного раздражения гораздо быстрее, чем зона разряда. Причем при любой интенсивности раздражения подпорогового возбуждения нейронов всегда больше, чем тех, что разряжаются, т.е. соответствуют импульсной активности (соотношение примерно 80:20).
Как в свете этих данных представить себе тонус нервных центров? Очевидно, что тонус центров определяется соотношением нейронов, которые «молчат», и нейронов, которые разряжаются, т.е. нейронов подпороговой зоны и зоны разряда. Если схематично изобразить нервный центр, который состоит из 50 нейронов, то тонус такого центра намного выше, когда импульсная активность наблюдается у 25 нейронах из 50, чем тогда, когда раздражаются только 10 клеток.
Можно допустить, что чем выше тоническая активность центра, т.е. чем больше нейронов генерирует потенциалы действия в данный момент, тем меньше возможности центра развивать рефлекторную деятельность в ответ на дополнительное раздражение. Центр слева находится в состоянии высокого тонуса, но у него только половина нейронов может «включиться» в ответ на дополнительные стимулы. Центр справа имеет низкую тоническую активность, но у него больше резервов для «включения» в рефлекторные реакции. Действительно, центры с постоянным тонусом (например, ядро блуждающего нерва) имеют тем меньшую рефлекторную возбудимость, чем выше их тоническая активность.
Нервные центры легко утомляются. Это проявляется постепенным снижением и даже полным прекращением импульсных разрядов при длительном раздражении афферентных волокон. В то же время раздражение эфферентного нерва (например, мышечного) еще продолжает вызывать сокращение мышцы. Если учесть, что нерв практически не устает, то усталость, которая развивается, прежде локализуется в нервном центре. Усталость центров связана главным образом с резким нарушением синаптической передачи (уменьшение запасов и синтеза медиатора, снижение чувствительности к медиатору постсинаптической мембраны, уменьшение энергетических резервов нервной клетки и др.).
Чувствительность нервных центров к гипоксии. Функции нервных центров зависят от снабжения их кислородом. Нуждаясь в большом количестве кислорода (мозг человека потребляет примерно 40-50 мл кислорода в 1 мин, т.е. 1/6-1/8 часть кислорода, необходимого организму в состоянии покоя), нервные клетки, особенно высших отделов ЦНС, очень чувствительны к его недостатку (гипоксии). Полное или частичное прекращение кровообращения мозга ведет к тяжелым нарушениям его деятельности и к гибели нервных клеток. Даже кратковременное резкое падение кровяного давления в мозгу вызывает у человека немедленную потерю сознания. Клетки коры большого мозга подлежат необратимым изменениям и погибают уже через 5-6 мин после полного прекращения кровообращения, при температуре 37 ° С функции клеток ствола головного мозга и спинного мозга нарушаются соответственно через 15 и 30 мин.
Нервные клетки и синапсы обладают избирательной чувствительностью к некоторым ядам, в частности к стрихнину, морфину, алкоголю, наркотическим веществам (эфир, хлороформ, барбитураты) и другие, их изучением занимается нейрофармакология.
Нервный центр — это совокупность нейронов, необходимых для осуществления определенного рефлекса или регуляции определенной функции.
Основными клеточными элементами нервного центра являются многочисленные , скопление которых формирует нервные ядра. В состав центра могут входить нейроны, рассеянные за пределами ядер. Нервный центр может быть представлен структурами мозга, располагающимися на нескольких уровнях центральной нервной системы (например, кровообращения, пищеварения).
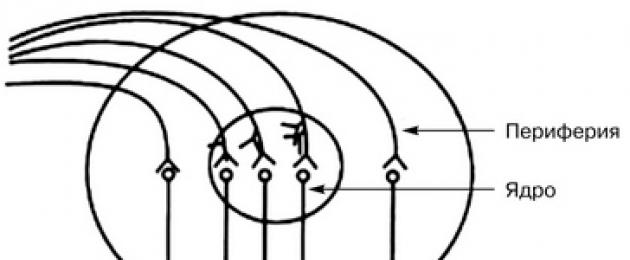
Любой нервный центр состоит из ядра и периферии.
Ядерная часть нервного центра представляет собой функциональное объединение нейронов, в которое поступает основная информация от афферентных путей. Повреждение этого участка нервного центра приводит к повреждению или существенному нарушению осуществления данной функции.
Периферическая часть нервного центра получает небольшую порцию афферентной информации, и ее повреждение вызывает ограничение или уменьшение объема выполняемой функции (рис. 1).
Функционирование центральной нервной системы осуществляется благодаря деятельности значительного числа нервных центров, представляющих собой ансамбли нервных клеток, объединенных с помощью синаптических контактов и отличающихся огромным разнообразием и сложностью внутренних и внешних связей.

Рис. 1. Схема общего строения нервного центра
В нервных центрах выделяют следующие иерархические отделы: рабочие, регуляторные и исполнительные (рис. 2).

Рис. 2. Схема иерархического подчинения разных отделов нервных центров
Рабочий отдел нервного центра ответствен за осуществление данной функции. Например, рабочий отдел дыхательного центра представлен центрами вдоха, выдоха и пневмотаксиса, расположенными в и варолиевом мосту; нарушение этого отдела вызывает остановку дыхания.
Регуляторный отдел нервного центра - это центр, расположенный в и регулирующий активность рабочего отдела нервного центра. В свою очередь, активность регуляторного отдела нервного центра зависит от состояния рабочего отдела, который получает афферентную информацию, и от внешних стимулов среды. Так, регуляторный отдел дыхательного центра расположен в лобной доле коры больших полушарий и позволяет произвольно регулировать легочную вентиляцию (глубину и частоту дыхания). Однако эта произвольная регуляция небезгранична и зависит от функциональной активности рабочего отдела, афферентной им пульсации, отражающей состояние внутренней среды (в данном случае рН крови, концентрации углекислого газа и кислорода в крови).
Исполнительный отдел нервного центра - это двигательный центр, расположенный в спинном мозге и передающий информацию от рабочего отдела нервного центра к рабочим органам. Исполнительный отдел дыхательного нервного центра расположен в передних рогах грудного отдела спинного мозга и транслирует приказы рабочего центра к дыхательным мышцам.
С другой стороны, одни и те же нейроны головного и спинного мозга могут участвовать в регуляции разных функций. Например, клетки центра глотания участвуют в регуляции не только акта глотания, но и акта рвоты. Этот центр обеспечивает все последовательные стадии акта глотания: движение мышц языка, сокращение мышц мягкого неба и его поднятие, последующее сокращение мышц глотки и пищевода при прохождении пищевого комка. Эти же нервные клетки обеспечивают сокращение мышц мягкого нёба и его поднятие во время акта рвоты. Следовательно, одни и те же нервные клетки входят и в центр глотания, и в центр рвоты.
Свойства нервных центров
Свойства нервных центров зависят от их строения и механизмов передачи возбуждения в . Выделяются следующие свойства нервных центров:
- Односторонность проведения возбуждения
- Синаптическая задержка
- Суммация возбуждения
- Трансформация ритма
- Утомляемость
- Конвергенция
- Дивергенция
- Иррадиация возбуждения
- Концентрация возбуждения
- Тонус
- Пластичность
- Облегчение
- Окклюзия
- Реверберация
- Пролонгирование
Одностороннее проведение возбуждение в нервном центре. Возбуждение в ЦНС проводится в одном направлении с аксона на дендрит или тело клетки следующего нейрона. Основу этого свойства составляют особенности морфологической связи между нейронами.
Одностороннее проведение возбуждения зависит от и гуморальной природы передачи в нем импульса: медиатор, осуществляющий передачу возбуждения, выделяется только в пресинаптическом окончании, а рецепторы, воспринимающие медиатор, расположены на постсинаптической мембране;
Замедление проведения возбуждения (центральная задержка). В системе рефлекторной дуги медленнее всего проводится возбуждение в синапсах ЦНС. В связи с этим центральное время рефлекса зависит от количества вставочных нейронов.
Чем сложнее рефлекторная реакция, тем больше центральное время рефлекса. Его величина связана со сравнительно медленным проведением возбуждения через последовательно включенные синапсы. Замедление проведения возбуждения создается вследствие относительной длительности осуществляющихся в синапсах процессов: выделения медиатора через пресинаптическую мембрану, его диффузии через синаптическую щель, возбуждения постсинаптической мембраны, возникновения возбуждающего постсинаптического потенциала и его перехода в потенциал действия;
Трансформация ритма возбуждения. Нервные центры способны изменять ритм поступающих к ним импульсов. Они могут отвечать на одиночные раздражители серией импульсов или на раздражители небольшой частоты — возникновением более частых потенциалов действия. В результате ЦНС посылает к рабочему органу количество импульсов, относительно независимое от частоты раздражений.
Это связано с тем, что нейрон является изолированной единицей нервной системы, к нему в каждый момент приходит множество раздражений. Под их влиянием происходит изменение мембранного потенциала клетки. Если создается небольшая, но продолжительная деполяризация (длительный возбуждающий постсинаптический потенциал), то один стимул вызывает серию импульсов (рис. 3);

Рис. 3. Схема трансформации ритма возбуждения
Последействие - способность сохранять возбуждение после окончания действия раздражителя, т.е. афферентных импульсов нет, а эфферентные продолжают действовать еще некоторое время.
Последействие объясняется наличием следовой деполяризации. Если следовая деполяризация длительна, то на ее фоне в течение нескольких миллисекунд могут возникать потенциалы действия (ритмическая активность нейрона), вследствие чего сохраняется ответная реакция. Но это дает сравнительно короткий эффект последействия.
Более длительное последействие связано с наличием кольцевых связей между нейронами. В них возбуждение как бы само себя поддерживает, возвращаясь по коллатералям к первоначально возбужденному нейрону (рис. 4);

Рис. 4. Схема кольцевых связей в нервном центре (по Лоренто де Но): 1 — афферентный путь; 2-промежуточные нейроны; 3 — эфферентный нейрон; 4 — эфферентный путь; 5 — возвратная ветвь аксона
Облегчение проведения или проторение пути. Установлено, что после возбуждения, возникшего в ответ на ритмическое раздражение, следующий стимул вызывает больший эффект, или для поддержания прежнего уровня ответной реакции требуется меньшая сила последующего раздражения. Это явление получило название «облегчение».
Его можно объяснить тем, что при первых стимулах ритмического раздражителя происходит перемещение пузырьков медиатора ближе к пресинаптической мембране и при последующем раздражении медиатор быстрее выделяется в синаптическую щель. Это, в свою очередь, приводит к тому, что вследствие суммации возбуждающего постсинаптического потенциала быстрее достигается критический уровень деполяризации и возникает распространяющийся потенциал действия (рис. 5);

Рис. 5. Схема облегчения проведения
Суммация, впервые описанная И.М. Сеченовым (1863) и заключающаяся в том, что слабые по силе раздражители, не вызывающие видимой реакции, при частом повторении могут суммироваться, создавать надпороговую силу и вызывать эффект возбуждения. Различают два вида суммации — последовательную и пространственную.
- Последовательная суммация в синапсах возникает в том случае, когда по одному и тому же афферентному пути к центрам поступает несколько подпороговых импульсов. В результате суммации местного возбуждения, вызванного каждым подпороговым стимулом, возникает ответная реакция.
- Пространственная суммация заключается в появлении рефлекторной реакции в ответ на два или несколько подпороговых стимулов, приходящих в нервный центр по разным афферентным путям (рис. 6);

Рис. 6. Свойство нервного центра — суммация пространственная (Б) и последовательная (А)
Пространственную суммацию, как и последовательную, можно объяснить тем, что при подпороговом раздражении, пришедшем по одному афферентному пути, выделяется недостаточное количество медиатора для того, чтобы вызвать деполяризацию мембраны до критического уровня. Если же импульсы приходят одновременно несколькими афферентными путями к одному и тому же нейрону, в синапсах выделяется достаточное количество медиатора, необходимое для пороговой деполяризации и возникновения потенциала действия;
Иррадиация. При возбуждении нервного центра нервные импульсы распространяются на соседние центры и приводят их в деятельное состояние. Это явление получило название иррадиации. Степень иррадиации зависит от количества вставочных нейронов, степени их миелинизации, силы раздражителя. Со временем в результате афферентной стимуляции только одного нервного центра зона иррадиации уменьшается, происходит переход к процессу концентрации, т.е. ограничению возбуждения только в одном нервном центре. Это является следствием уменьшения синтеза медиаторов во вставочных нейронах, в результате чего биотоки не передаются из данного нервного центра на соседние (рис. 7 и 8).

Рис. 7. Процесс иррадиации возбуждения в нервных центрах: 1, 2, 3 — нервные центры

Рис. 8. Процесс концентрации возбуждения в нервном центре
Выражением данного процесса является точная координированная двигательная реакция в ответ на раздражение рецептивного поля. Формирование любых навыков (трудовых, спортивных и т.д.) обусловлено тренировкой двигательных центров, основу которых составляет переход от процесса иррадиации к концентрации;
Индукция. Основой взаимосвязи между нервными центрами является процесс индукции — наведение (индуцирование) противоположного процесса. Сильный процесс возбуждения в нервном центре вызывает (наводит) торможение в соседних нервных центрах (пространственная отрицательная индукция), а сильный тормозной процесс индуцирует в соседних нервных центрах возбуждение (пространственная положительная индукция). При смене этих процессов в пределах одного центра говорят о последовательной отрицательной или положительной индукции. Индукция ограничивает распространение (иррадиацию) нервных процессов и обеспечивает концентрацию. Способность к индукции в значительной степени зависит от функционирования тормозных вставочных нейронов — клеток Реншоу.
От степени развития индукции зависят подвижность нервных процессов, возможность выполнения движений скоростного характера, требующих быстрой смены возбуждения и торможения.
Индукция является основой доминанты — образования нервного центра повышенной возбудимости. Это явление впервые было описано А.А. Ухтомским. Доминантный нервный центр подчиняет себе более слабые нервные центры, притягивает их энергию и за счет этого еще более усиливается. В результате этого раздражение различных рецепторных полей начинает вызывать рефлекторный ответ, характерный для деятельности этого доминантного центра. Доминантный очаг в ЦНС может возникать под влиянием разных факторов, в частности сильной афферентной стимуляции, гормональных воздействий, мотиваций и т.д. (рис. 9);
Дивергенция и конвергенция. Способность нейрона устанавливать многочисленные синаптические связи с различными нервными клетками в пределах одного или разных нервных центров называется дивергенциеи. Например, центральные окончания аксонов первичного афферентного нейрона образуют синапсы на многих вставочных нейронах. Благодаря этому одна и та же нервная клетка может участвовать в различных нервных реакциях и контролировать большое число других , что приводит к иррадиации возбуждения.

Рис. 9. Формирование доминанты за счет пространственной отрицательной индукции
Схождение различных путей проведения нервных импульсов к одному и тому же нейрону получило название конвергенции. Простейшим примером конвергенции является замыкание на одном двигательном нейроне импульсов от нескольких афферентных (чувствительных) нейронов. В ЦНС большинство нейронов получают информацию от разных источников благодаря конвергенции. Это обеспечивает пространственную суммацию импульсов и усиление конечного эффекта (рис. 10).

Рис. 10. Дивергенция и конвергенция
Явление конвергенции было описано Ч. Шеррингтоном и получило название воронки Шеррингтона, или эффекта общего конечного пути. Данный принцип показывает, как при активации различных нервных структур формируется конечная реакция, что имеет первостепенное значение для анализа рефлекторной деятельности;
Окклюзия и облегчение. В зависимости от взаимного расположения ядерных и периферических зон разных нервных центров может проявиться при взаимодействии рефлексов явление окклюзии (закупорки) или облегчения (суммации) (рис. 11).

Рис. 11. Окклюзия и облегчение
Если происходит взаимное перекрывание ядер двух нервных центров, то при раздражении афферентного поля первого нервного центра условно возникают два двигательных ответа. При активации только второго центра также возни каст два двигательных ответа. Однако при одновременной стимуляции обоих центров суммарный двигательный ответ равен только трем единицам, а не четырем. Это обусловлено тем, что один и тот же мотонейрон относится одновременно к обоим нервным центрам.
Если происходит перекрывание периферических отделов разных нервных центров, то при раздражении одного центра возникает одна ответная реакция, то же наблюдается и при раздражении второго центра. При одновременном возбуждении двух нервных центров возникает три ответных реакции. Потому что мотонейроны, находящиеся в зоне перекрывания и не дающие ответа при изолированном раздражении нервных центров, получают при одновременной стимуляции обоих центров суммарную дозу медиатора, что приводит к пороговому уровню деполяризации;
Утомляемость нервного центра. Нервный центр обладает малой лабильностью. Он постоянно получает от множества высоколабильных нервных волокон большое количество стимулов, превышающих его лабильность. Поэтому нервный центр работает с максимальной загрузкой и легко утомляется.
Исходя из синаптических механизмов передачи возбуждения утомление в нервных центрах может объясняться тем, что но мере работы нейрона истощаются запасы медиатора и становится невозможной передача импульсов в синапсах. Кроме того, в процессе деятельности нейрона наступает постепенное снижение чувствительности его рецепторов к медиатору, что называется десенситизацией;
Чувствительность нервных центров к кислороду и некоторым фармакологическим веществам. В нервных клетках осуществляется интенсивный обмен веществ, для чего необходимы энергия и постоянный приток нужного количества кислорода.
Особенно чувствительны к недостатку кислорода нервные клетки коры больших полушарий головного мозга, после пяти-шести минут кислородного голодания они погибают. У человека даже кратковременное ограничение мозгового кровообращения приводит к потере сознания. Недостаточное снабжение кислородом легче переносят нервные клетки мозгового ствола, их функция восстанавливается через 15-20 мин после полного прекращения кровоснабжения. А функция клеток спинного мозга восстанавливаются даже после 30 мин отсутствия кровообращения.
По сравнению с нервным центром нервное волокно малочувствительно к недостатку кислорода. Помешенное в атмосферу азота, оно только через 1,5 ч прекращает проведение возбуждения.
Нервные центры обладают специфической реакцией на различные фармакологические вещества, что свидетельствует об их специфичности и своеобразии протекающих в них процессов. Например, никотин, мускарин блокируют проведение импульсов в возбуждающих синапсах; их действие приводит к падению возбудимости, уменьшению двигательной активности и полному ее прекращению. Стрихнин, столбнячный токсин выключают тормозящие синапсы, что приводит к повышению возбудимости ЦНС и увеличению двигательной активности вплоть до общих судорог. Некоторые вещества блокируют проведение возбуждения в нервных окончаниях: кураре — в концевой пластинке; атропин — в окончаниях парасимпатической нервной системы. Есть вещества, действующие на определенные центры: апоморфин — на рвотный; лобелии — на дыхательный; кардиазол — на двигательную зону коры; мескалин — на зрительные центры коры и др.;
Пластичность нервных центров. Под пластичностью понимают функциональную изменчивость и приспособляемость нервных центров. Это особенно ярко проявляется при удалении разных отделов мозга. Нарушенная функция может восстанавливаться, если были частично удалены какие-то отделы мозжечка или коры больших полушарий. О возможности полной перестройки центров свидетельствуют опыты по сшиванию функционально различных нервов. Если перерезать двигательный нерв, иннервирующий мышцы конечностей, и его периферический конец сшить с центральным концом перерезанного блуждающего нерва, регулирующего внутренние органы, то через некоторое время периферические волокна двигательного нерва перерождаются (вследствие их отделения от тела клетки), а волокна блуждающего нерва прорастают к мышце. Последние образуют в мышце синапсы, свойственные соматическому нерву, что приводит к постепенному восстановлению двигательной функции. В первое время после восстановления иннервации конечности раздражение кожи вызывает свойственную блуждающему нерву реакцию — рвоту, гак как возбуждение от кожи по блуждающему нерву поступает в соответствующие центры продолговатого мозга. Через некоторое время раздражение кожи начинает вызывать обычную двигательную реакцию, поскольку происходит полная перестройка деятельности центра.
Процессы обработки информации, поступающей в нервный центр (если он сенсорный), или формирование команд к исполнительным органам (в ефекторному центре) обусловлены взаимодействием нейронов посредством синаптических контактов. В таком случае можно обнаружить явления, что называют дивергенцией и конвергенцией (рис. 37).
Дивергенция - это способность нейрона устанавливать многочисленные связи с другими нейронами. Вследствие этого одна и та же клетка может участвовать в различных нервных процессах и реакциях, контролировать большое число других нейронов, то есть каждый нейрон может обеспечить распространение импульсов - иррадиацию возбуждения. Процессы дивергенции более типичны для афферентных отделов ЦНС.
Конвергенция - схождение различных путей проведения нейрон-ных импульсов к одной и той же нервной клетки, больше присуща нервным центрам эфферентных отделов.
Большинство нервных центров представлено скоплением разнообразных нейронов. Среди них бывают как возбуждающие, так и тормозные нейроны, нейроны сенсорные и моторные (афферентные или эфферентные). их довольно сложное взаимодействие и обеспечивает выполнение соответствующих функций.
Взаимодействие рефлексов
В процесс регуляции большинства сложных функций организма, организации рефлекторного ответа очень часто привлекаются несколько нервных центров, которые могут размещаться даже на различных этажах ЦНС. Обусловлено это філогенетичними особенностями формирования ЦНС. Появление "младшего" отдела сопровождалась формированием в нем новых центров регуляции. Но и "старые" нервные центры, расположенные в низших отделах, сохраняли свойственные им функции. При этом терялась абсолютная автономность отдельных сегментов ЦНС, все большая часть функций "переходила" высшим отделам. Этот процесс получил название енцефалізації функций. Поскольку головной мозг формировался поэтапно, от заднего мозга до переднего с его большими полушариями, то с формированием коры больших полушарий происходит подчинение ей других отделов ЦНС, то есть кортикалізація функций.
Поскольку каждый из нервных центров отвечает за определенные рефлексы, во время их взаимодействия можно говорить о взаимодействии различных рефлексов. Это взаимодействие осуществляется на основе определенных закономерностей, которые позволяют ЦНС решать свои функциональные задачи как с целенаправленного регулирования различных систем организма, так и организации его поведения в конкретных, постоянно меняющихся условиях внешней среды.
Можно выделить такие принципы координации функций ЦНС.
1. Торможение в ЦНС.
Важной частью нейронных цепей, образующих рефлекторные дуги, является наличие тормозных нейронов (рис. 38). Вследствие этого ослабляется или совсем прекращается интенсивный процесс возбуждения, что в основном обеспечивает упорядочение проявления рефлекса. Пример торможения - реципрокне торможения мышц-антагонистов на уровне мотонейронов спинного мозга (рис. 38, а). Процесс тормозного влияния запускается через специальные тормозные клетки Реншо, содержащихся в спинном мозге. При поступлении афе
Рис. 38. Торможение в ЦНС : а - участие тормозных интернейронов спинного мозга (Г) в регуляции деятельности мышц-антагонистов: торможение (-) мотонейрона мышцы-разгибателя (МР) во время возбуждения (+) мотонейрона мышцы-сгибателя (МЗ); б - поворотное (постсинаптичне) торможения (МН - мотонейрон, Г - тормозная клетка Реншо; М - мышца); в - торможения нейронов промежуточного мозга с участием тормозной корзинного клетки (Г); г - пресинаптичне торможения (Г - тормозная клетка; Н - нейрон; Пр - пресинаптичне волокно; за Екклсом)
рентной импульсации они активируются одновременно с нейронами, которые возбуждаются, обеспечивая реципрокный взаимосвязь при осуществлении двигательных рефлексов: мотонейроны одних мышц возбуждаются, а их антагонистов - тормозятся.
Второй, довольно распространенный, тип первичного торможения - возвратное торможение (рис. 38, б). Клетки Реншо располагаются еще и таким образом, что через коллатерали возбужденного мотонейрона вызывают его торможение. Это типичный пример отрицательной обратной связи, когда подавляется чрезмерная импульсация.
2. Иррадиация и концентрация нервных процессов.
Возбуждение, возникшее в одном из центров, может распространяться через коллатерали и синапсы на другие центры. Процесс иррадиации чаще всего развивается в случае действия сильного раздражителя. Например, во время сильного давления на лапку лягушки сокращаются не одна, а все конечности. Через некоторое время иррадиация меняется на явление концентрации возбуждения в необходимом центре. Это обусловлено действием тормозных синаптических связей. Процессы иррадиации и концентрации основываются на свойствах конвергенции и дивергенции.
3. Явления суммации и окклюзии
(рис. 39). Суммация (облегчение) оказывается во время воздействия нескольких подпороговых стимулов (с разных рецепторов), каждый из которых, действуя отдельно, не вызывает ответа. А их суммация (при условии рядом расположенных синаптических полей) способствует проявлению ответа нервного центра (явление облегчения).

Рис. 39. Схема, иллюстрирующая явление облегчения (1) и окклюзии (2) нервного импульса: а - в центральных кругах изображены нейроны, которые возбуждаются как при изолированной, так и одновременного раздражения нервных волокон (В, 2); пунктирными линиями обведены нейроны, которые возбуждаются только за одновременного раздражения обеих нервных волокон; б -в центральной части, образованной кругами, перекрещивающихся расположены нейроны, которые возбуждаются как при изолированной, так и одновременного раздражения нервных волокон
Противоположное явление - окклюзии (заклинивания) - развивается при тех же условиях расположения синаптических полей, но при одновременном действии нескольких раздражителей надграничної силы. Суммарная ответ может быть меньшим, чем арифметическая сумма ответов на каждый из раздражителей в отдельности, что происходит за "перекрытия" как на уровне рецептора, так и общих центральных нейронов.
4. Принцип "общего конечного пути"
(рис. 40). Он основывается на явлении конвергенции. Афферентных входов в ЦНС значительно больше, чем эфферентных выходов. Следовательно, один и тот же рефлекс можно вызвать, раздражая различные рефлекторные поля.
5. Принцип доминантного очага.
Содержание принципа заключается в том, что в случае одновременного возбуждения нескольких нервных центров один из очагов может стать доминантным. Вследствие этого к нему могут активно привлекаться (иррадиировать) возбуждения из других очагов, что приведет к суммации возбуждения, усиливая доминантное возбуждение. Высокую возбудимость нейронов обусловливают соответствующая аферентна импульсация (например из переполненного мочевого пузыря), гуморальные влияния. В результате оказывается, что для организма функция этого центра в конкретный временной промежуток становится важнейшей.
Основные признаки доминантного очага следующие:
1) стойкость возбуждения во времени;
2) повышенная возбудимость;
3) способность к суммации. Доминанта - это физиологическое основание возникновения взаимосвязей между отдельными нервными центрами при формировании условных рефлексов, основа внимания.

Рис. 40. а - клетки спинномозговых
ганглий; б - промежуточные нейроны; в - мотонейроны; г - мышцу (зачеркнуто тела нейронов, которые тормозят нервные импульсы; за Шеррингтоном)
Исходя из физиологических представлений, нервный «центр» может располагаться на разных уровнях ЦНС и участвовать в регуляции какой-либо фиpиологической функции (дыхание, пищеварение и т.д.) или в совершении какого-либо рефлекса.
К функциональным свойствам рефлекторных центров относятся: иррадиация
возбуждения; конвергенция
идивергенция
; суммирование; синаптическое облегчение и окклюзия; трансформация ритма, реверберация возбуждения; тоническое состояние центров, их быстрая утомляемость, большая чувствительность к недостатку кислорода и к действию некоторых ядов.
Иррадиация возбуждения
Активное распространение возбуждения в ЦНС, особенно при сильном и длительном раздражении, получило название иррадиации. Возможность иррадиации в ЦНС обусловлена наличием в ней многочисленных ответвлений отростков (аксонов, дендритов) нервных клеток и цепей интернейронов, которые соединяют между собой различные нервные центры (благодаря этому возбуждение распространяется определенными путями и с определенной последовательностью). Важную роль в иррадиации возбуждения в структурах мозга играет ретикулярная формация.
Усиление раздражения или повышение возбудимости ЦНС сопровождается усилением иррадиации возбуждения в ней. Тормозные нейроны и синапсы препятствуют иррадиации возбуждения или ограничивают ее. При введении стрихнина, блокирующего постсинаптическое торможение, возникает сильное возбуждение ЦНС, которое сопровождается судорогами всех скелетных мышц. Иррадиация может стать патологической в связи с возникновением сильного очага возбуждения и с изменением свойств нервной ткани, усиливает распространение возбуждения. Это бывает при эпилепсии.
Конвергенция возбуждения
На каждом из нейронов ЦНС конвергирует (сходятся) различные афферентные волокна. Таких афферентных входов для большинства нейронов много десятков и даже тысяч. Так, на мотонейронах заканчиваются в среднем 6000 коллатералей аксонов, которые поступают от периферических рецепторов и различных структур мозга, образуя возбуждающие и тормозные синапсы. Это такое универсальное явление, можно говорить о принципе конвергенции в нейронах и их связях. Благодаря этому явлению в один и тот же нейрон одновременно поступают многочисленные и разнообразные потоки возбуждений, которые затем подлежат сложной обработке и перекодируются и формируются в единое возбуждение - аксонноу, что идет к следующему звену нервной сетки. Конвергенция возбуждения на нейроне является универсальным фактором его интегративной деятельности.
Различают мультисенсорную, мультибиологическую и сенсорно-биологическую формы конвергенции. В первом случае на нейрон поступают сигналы различной сенсорной модальности (зрительные, слуховые, болевые и др.), во втором - потоки возбуждений различной биологической модальности (пищевые, половые и др.), в третьем - сигнализация (зрительная, пищевая) и другие.
Дивергенция возбуждения
Дивергенция (расхождение) возбуждения - способность одиночного нейрона устанавливать в многочисленных синаптических связях с различными нервными клетками. Например, афферентные волокна периферических рецепторов, входя в спинной мозг в составе задних корешков, дальше разветвляются на многочисленные коллатерали, которые идут к спинальным нейронам. Благодаря дивергенции одна и та же нервная клетка может принимать участие в организации различных реакций и контролировать большое количество нейронов. Одновременно каждый нейрон может обеспечивать широкое перераспределение импульсов, что ведет к иррадиации возбуждения. Конвергенция и дивергенция взаимно связаны.
Реверберация возбуждения
Циркуляция возбуждения замкнутыми нейронами и их цепями в ЦНС называется реверберацией. Возбуждение одного из нейронов, входящих в эту цепь, передается на другой (или другие), а коллатералям аксонов снова возвращается к нервной клетки и т.д.
Реверберация возбуждения наблюдается в так называемом рефлекторном последействии, когда рефлекторный акт заканчивается не сразу после прекращения, а через некоторый (иногда длительный) период, а также играет определенную роль в механизмах кратковременной (оперативной) памяти. Сюда же относится корково-подкорковая реверберация, которая играет важную роль в высшей нервной деятельности (поведении) человека и животных.
Лат. diverqere - направляется в разные стороны - способность одиночного нейрона устанавливать многочисленные синаптические связи с различными нервными клетками. Благодаря процессу дивергенции одна и та же клетка может участвовать в организации различных реакций и контролировать большее число нейронов. В то же время каждый нейрон может обеспечивать широкое перераспределение импульсов, что приводит к иррадиации возбуждения.
Иррадиация (от лат. irradio - сияю, испускаю лучи) в физиологии, распространение процесса возбуждения или торможения в центральной нервной системе. Важную роль И. играет в деятельности коры больших полушарий головного мозга. И. возбуждения особенно отчётливо проявляется при сильном раздражении, когда в рефлекторный ответ вовлекаются нервные центры, обычно в нём не участвующие. Так, на умеренное болевое раздражение кожи стопы животное отвечает сгибанием лапы в голеностопном суставе; увеличение силы раздражения приводит к сгибанию ноги в коленном и тазобедренном суставах. При изучении действия тормозного условного раздражителя И. П. Павловым было показано, что торможение также может распространяться (иррадиировать) в клетках коры больших полушарий
Принцип иррадиации , или дивергенции, возбуждения ЦНС. Иррадиация возбуждения (от лат. irradio, озарять, освещать) - это распространение процесса возбуждения из одного участка ЦНС в другой. Каждый нейрон за счет многочисленных ответвлений (дивергенции), заканчивающихся синапсами, и большого числа вставочных нейронов связан со многими другими нейронами. Поэтому нервные импульсы от одного нейрона могут быть направлены к тысяче других нейронов. Процесс иррадиации возбуждения регулируется различными механизмами. Он может быть усилен, например, за счет активации ретикулярной формации ствола мозга. С другой стороны, процесс иррадиации ограничивается, и это важное действие осуществляется с участием многочисленных тормозных нейронов. Например, в спинном мозге ограничение иррадиации осуществляется благодаря механизму возвратного торможения с помощью специальных тормозных интернейронов - клеток Реншоу. Иррадиация возбуждения играет исключительно важную роль, так как позволяет обмениваться многочисленными потоками информации различным структурам мозга. Именно за счет иррадиации происходит обмен информацией между первой и второй сигнальными системами (элективная иррадиация), что существенно увеличивает возможности высшей нервной деятельности человека. Благодаря иррадиации возбуждения осуществляется интеграция деятельности правого и левого полушарий.
Принцип концентрации возбуждения
Возбуждение и торможение могут либо иррадиировать, либо концентрироваться. Концентрация - это явление, противоположное иррадиации. Оно возникает в тот момент, когда иррадиация достигает определенной границы, после чего распространение возбуждения или торможения идет в обратном направлении, т. е. в исходный пункт. Если нейроны находятся в заторможенном состоянии, это препятствует распространению на них процесса возбуждения, и наоборот, если они находятся в состоянии возбуждения, торможением они охватываются труднее. На скорость иррадиации и концентрации нервных процессов оказывает влияние и тип нервной системы, ее индивидуальные особенности.
Принцип индукции нервных процессов. Индукция отражает проявление процессов иррадиации возбуждения и торможения в коре больших полушарий. Принято считать, что индукция - (от лат. Inductio - введение, наведение) обозначает возникновение нервного процесса, противоположного по знаку процессу, вызванному условным раздражителем (положительным или отрицательным, т. е. тормозным). Одновременная индукция заключается в том, что формирование в каком-либо центре коры больших полушарий концентрированного возбуждения вызывает в прилежащих к этому центру зонах торможение (отрицательная одновременная индукция), а такое же концентрированное торможение вызывает в этих зонах возбуждение (положительная одновременная индукция). Таким образом, при одновременной индукции нервный процесс вызывает в другом участке коры процесс, противоположный по знаку (процесс возбуждения вызывает процесс торможения и наоборот), а при последовательной индукции происходит смена противоположных нервных процессов в одном и том же участке.
Принцип конвергенции возбуждения (или принцип общего конечного пути, воронка Шеррингтона). Конвергенция нервных импульсов (от лат. convergo, convergere - сближать, сходиться) означает схождение к одному нейрону двух или нескольких различных возбуждений одновременно. Это явление было открыто Ч. Шеррингтоном. Он показал, что одно и то же движение, например рефлекторное сгибание конечности в коленном суставе, можно вызвать путем раздражения различных рефлексогенных зон. В связи с этим им было введено понятие "общего конечного пути", или "принципа воронки", согласно которому потоки импульсов от различных нейронов могут сходиться на одном и том же нейроне (в данном случае - на альфа-мотонейронах спинного мозга). В частности, Ч. Шеррингтон обнаружил схождение к одним и тем же промежуточным или эфферентным нейронам различных афферентов от разных участков общего рецептивного поля (в спинном и продолговатом мозге) или даже от разных рецептивных полей (в высших отделах головного мозга). В настоящее время показано, что конвергенция возбуждения, так же как и дивергенция возбуждения, - очень распространенное явление в ЦНС. Основой для конвергенции (как и для иррадиации) является определенная морфологическая и функциональная структура различных отделов мозга. Очевидно, что часть конвергентных путей является врожденными, а другая часть (главным образом в коре большого мозга) - приобретенная в результате обучения в процессе онтогенеза. Формирование новых конвергентных отношений для нейронов коры большого мозга в процессе онтогенеза во многом связаны с формированием в коре доминантных очагов возбуждения, которые способны "притягивать" к себе возбуждение от других нейронов.
Принцип доминанты, или господствующего очага возбуждения. Доминанта (от лат. dominans, dominantis - господствующий) - это временно господствующая рефлекторная система, обусловливающая интегральный характер функционирования нервных центров в какой-либо период времени и определяющая целесообразное поведение животного и человека. В целом принцип доминанты означает, что текущая деятельность мозга определяется наличием господствующего (доминантного) очага возбуждения, или господствующего нейронного объединения, которое в данный момент времени подавляет и подчиняет себе деятельность остальных нейронных образований. Таким образом, благодаря формированию доминантного очага (доминантного нейронного объединения) деятельность мозга организуется таким образом, чтобы удовлетворить потребность организма, наличие которой и сформировало доминантный очаг возбуждения. Детальное изучение свойств доминантного очага показало, что для совокупности нейронов, входящих в его состав, характерны повышенная возбудимость, повышенная стойкость возбуждения, или инертность (заключающаяся в том, что у таких нейронов сложно вызвать торможение), повышенная способность к суммированию возбуждения, высокая способность "притягивать" возбуждение от других нервных центров (и тем самым повышать свою активность). Доминанта, как один из основных принципов координационной деятельности ЦНС, имеет важное значение в жизни человека. Например, именно благодаря доминанте возможно сосредоточение психической деятельности (внимание) и выполнение умственной или физической трудовой деятельности (в данном случае - это трудовая доминанта). В период поиска пищи и ее поедания реализуется пищевая доминанта. В настоящее время в отношении человека выделяют различные виды доминант (пищевую, оборонительную, половую, игровую, трудовую и др.).
Принцип субординации , или соподчинения, также относится к категории важнейших принципов организации работы мозга. Согласно этому принципу, деятельность нижележащих отделов мозга контролируется и управляется вышележащими отделами ЦНС. Например, в двигательных системах мозга и в вегетативной нервной системе имеются нейронные объединения (нервные центры), расположенные в спинном мозге или в стволе мозга, которые подчиняются деятельности нейронных объединений (нервных центров), находящихся в гипоталамусе, таламусе, мозжечке, базальных ядрах и коре больших полушарий.
Принцип обратной связи (обратной афферентации) и копий эфферентаций . Согласно этому принципу, для точной координации деятельности различных нейронных объединений (нервных центров, рефлекторных дуг) необходима оптимальная по объему информация о результатах действия. Она поступает в мозг по сенсорным каналам. Отсутствие такой информации приводит к дезинтеграции деятельности мозга. Особенно наглядна роль обратной афферентации при реализации двигательной активности - нарушение проприоцептивной чувствительности, как правило, препятствует выполнению точных движений, а также нарушает возможность формирования и сохранения адекватной для данного движения позы.
Принцип реципрокности (сопряжения) возбуждения и торможения на уровне спинного мозга реализуется с участием реципрокного торможения, благодаря которому возникают безусловные двигательные реципрокные рефлексы. Реципрокное торможение осуществляется по механизму постсинаптического торможения, которое возникает с участием специальных вставочных тормозных нейронов.
Принципы кодирования информации в нервной системе . В целом вся информация или значительная ее часть, передаваемая в ЦНС от одного отдела к другому, заключена в пространственном и временном распределении импульсных потоков, при этом используются различные нейронные коды. Выделяют три основные группы кодов. Неимпульсные сигналы , для которых характерны внутри- и внеклеточные факторы. К внутриклеточным факторам относятся амплитудные характеристики рецепторных и синаптических потенциалов, амплитудные и пространственные характеристики изменений синаптической проводимости, пространственное и временное распределение характеристик мембранного потенциала и градуальные потенциалы в аксонных терминалях. Внеклеточные факторы - это высвобождение медиаторов и ионов калия, нейросекреция, электротонические взаимодействия. Импульсные сигналы в одиночных нейронах. Для импульсных кодов главными кандидатами являются коды пространственные ("меченые линии", т. е. представление информации номером канала) и временные - различные виды частотных или интервальных кодов (взвешенное среднее значение частоты, мгновенное значение частоты, частота разряда, форма интервальных гистограмм и т. д.). Выделяют также микроструктурное кодирование (временный узор импульсов), латентный код (момент появления или фазовые изменения разряда), числовой код (количество импульсов в пачке), код длинной пачки (длительность импульсации), наличие отдельного импульса или его отсутствие) изменение скорости распространения возбуждения в аксоне и пространственную последовательность явлений в аксоне. Ансамблевая активность (кодирование по ансамблю). В большинстве случаев в ЦНС используется пространственно-временное кодирование, когда информация о признаках сигнала передается канально и уточняется различными модификациями временных кодов.
2) Надпочечники (glandula suprarenalis, ед. ч.)-парные железы внутренней секреции, расположенные над верхними полюсами почек. У человека они находятся на уровне XI грудного - I поясничного позвонков, забрюшинно. Правый надпочечник имеет треугольную форму, левый - полулунную; вогнутые основания надпочечников примыкают к выпуклым полюсам почек. Вместе с почками надпочечники заключены в жировую капсулу (сapsula adiposa) и покрыты почечной фасцией (fascia renalis). Длина надпочечника взрослого человека варьирует от 30 до 70 мм, ширина - от 20 до 35 мм, толщина - от 3 до 10 мм, масса обоих надпочечников составляет 10 -14 г. Снаружи надпочечник покрыт соединительнотканной капсулой, от которой в паренхиму отходят перегородки, заключающие в себе сосуды и нервы и делящие паренхиму надпочечников на группы клеток и клеточные тяжи. В надпочечниках различают наружное корковое вещество, составляющее примерно 2/3 всей массы надпочечника, и внутреннее мозговое вещество. У новорожденных масса надпочечника в среднем 3,5 г. С возрастом она увеличивается, постепенно разграничиваются корковое и мозговое вещество, отдельные зоны коркового вещества.
Кровоснабжение надпочечников осуществляют три группы надпочечниковых артерий: верхняя, средняя, нижняя, проникающие в паренхиму в виде многочисленных капилляров, которые широко анастамозируют между собой и образуют в мозговом веществе синусоиды - расширения. Отток крови от надпочечников происходит через центральную и многочисленные поверхностные вены, впадающие в венозную сеть окружающих органов и тканей. Параллельно кровеносным расположены лимфатические капилляры, отводящие лимфу. Иннервируются надпочечники симпатическими (преимущественно) и парасимпатическими волокнами чревного, блуждающего и диафрагмального нервов. Корковое вещество надпочечника делится на клубочковую, пучковую и сетчатую зоны. Клубочковая зона, прилегающая тонким слоем к соединительнотканной капсуле, состоит из клеток неправильной формы. В пучковой, средней, наиболее широкой зоне группируются железистые клетки, расположенные радиальными колонками, а в сетчатой, внутренней, зоне группы клеток имеют вид неправильной сетки. Корковое вещество надпочечника богато липидами, окрашивающими его в желтый цвет. Гормоны коркового вещества кортикостероиды (глюко- и минералокортикоиды) синтезируются в митохондриях секреторных клеток из холестерина. Многообразное влияние кортикостероидов на все виды обмена веществ, сосудистый тонус, иммунитет и др. делает корковое вещество надпочечников важнейшим участком жизнеобеспечения человека в обычных условиях и в условиях адаптации к различным стрессам. В клубочковой зоне коркового вещества синтезируется альдостерон - основной минералокортикоид, участвующий в регуляции водно-солевого обмена. В пучковой зоне синтезируется преимущественно кортизол - глюкокортикоид, влияющий на белковый, жировой и углеводный обмен и на обмен нуклеиновых кислот, и кортикостерон, обладающий свойствами глюко- и минералокортикоида. В сетчатой зоне образуются половые гормоны, главным образом андрогены. Синтез кортикостероидов, прежде всего глюкокортикостероидов, регулируются адренокортикотропным гормоном. В глубине надпочечника находится мозговое вещество. Железистые клетки мозгового вещества получили название хромаффинных, или феохромных, т.к. избирательно окрашиваются солями хрома в желто-бурый цвет. Помимо железистых клеток в мозговом веществе надпочечника много нервных волокон и нервных клеток. Скопления хромаффинных нервных клеток, так называемых параганглиев, обнаруживают также по ходу легочного ствола и восходящей части аорты, в средостении находится поясничный аортальный параганглий и другие. Хромаффинные клетки секретируют три гормона (адреналин, норадреналин и дофамин), объединенных под общим названием катехоламины. Биосинтетическими предшественниками этих гормонов является аминокислота тирозин. Адреналин синтезируется только в надпочечниках; норадреналин и дофамин образуются также в параганглиях и многочисленных нейронах симпатической нервной системы. Все ткани, продуцирующие катехоламины, составляют адреналиновую систему. Активность хромаффинных клеток стимулируют различные воздействия окружающей и внутренней среды: эмоции, артериальная гипотензия, гипогликемия, физическая нагрузка, охлаждение и др.